
Discover
Researchers Reveal the Secrete of Nonrandom DNA Seg-regation in Human Cells
ProMyelocytic Leukemia (PML) protein controls various biological functions, such as apoptosis, senescence or stem cell self-renewal. PML may elicit these functions by scaffolding the spherical shells of PML Nuclear Bodies (NBs), which subsequently act as hubs of post-translational modifications, in particular sumoylation, for the broad range of proteins trafficking through their inner cores. PML NBs are disrupted in Acute Promyelocytic Leukemia (APL) driven by the PML-RARA oncogenic fusion protein. Arsenic trioxide (ATO) cures 70% APL patients through driving PML-RARA degradation and NB reformation. In non-APL cells, arsenic binding onto PML also amplifies NB formation. Yet, the actual molecular mechanism involved remain elusive.
The research team led by Professors Naranmandura Hua, ZHOU Chun from Zhejiang University School of Medicine, and Hugues De The,Valerie Lallemand-Breitenbach from France has made a significant breakthrough in understanding how arsenic targets the PML protein. The research team has successfully elucidate the crystal structure of PML B-box 2 (B2) and identify an α-helix that mediates B2 trimerization through hydrophobic interactions. Importantly, the α-helix-mediated trimer organizes a triad of free C213 cysteines to which trivalent arsenic covalently docks. This triggers ATO-induced PML-RARA-degradation, which is the first step in ATO's anti-leukemia cure. Their findings are detailed in an article titled “Structural Basis of PML/RARA Oncoprotein Targeting by Arsenic Unravels a Cysteine Rheostat Controlling PML Body Assembly and Function”, published in Cancer Discovery.
B-box domains are frequently present in TRIpartite Motif (TRIM) proteins. TRIM5α B2 was proposed to trimerize through a specific α-helix around a tryptophan core in vitro. We thus looked whether PML B2 monomers could self-assemble into a trimer through Alphafold2 molecular modeling. In the model, the trimer is predicted to be driven by α1-helix mediated interactions, involving hydrophobic interfaces and in particular, I202, C213, A216, L217 and L218. Supporting this notion, point mutations of these potentially interfacing residues (I202A, L218G, L217G, A216V) disrupted NB assembly and exhibited highly dynamic exchange with liquid-like behaviors, as analyzed by immunofluorescence and FRAP. Furthermore, SEC-MALS analysis revealed three forms of purified MBP-B2 with molecular weights corresponding to monomer, dimer, and critically, abundant trimer.
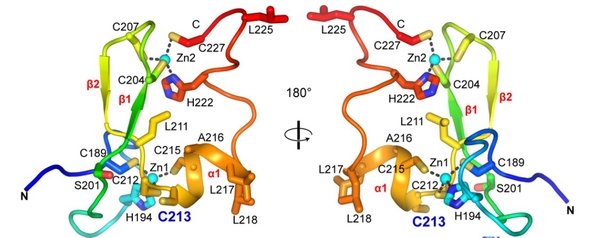
Figure 1. Crystal structure of PML B2 monomer.
ATO and other therapeutic arsenic derivatives provide trivalent arsenic (As(III) oxidation state) at physiological pH and in vivo. As(III) can form stable complexes with thiol groups through covalent binding to sulfur and may form complex with three glutathiones in solution. Critically, the key C213 residues are organized in a triad positioned at the center of the PML B2 helix trimer, fitting with an ideal arsenic docking. We thus hypothesized that ATO could targets PML NBs by interacting with this triad of C213 residues within the B2 trimer. To interrogate the relevance of this model in cellulo, we generated several point mutations. While mutations of C213 removing thiol group (C213S or C213A) and increasing steric hindrance (C213L or C213F) all disrupted basal PML NB assembly, replacing C213 by valine unexpectedly fully rescued the basal NB formation and normal dynamics. Strikingly, both PML C213V localization and dynamics were insensitive to ATO, implying that the sensor function was lost. Accordingly, alkylation by NEM, which disrupted WT NBs, had no effect on PMLC213V NBs, stressing the unique features of this cysteine residue. Altogether, this suggests that, within the B2 domain, C213 may directly contribute to arsenic binding.
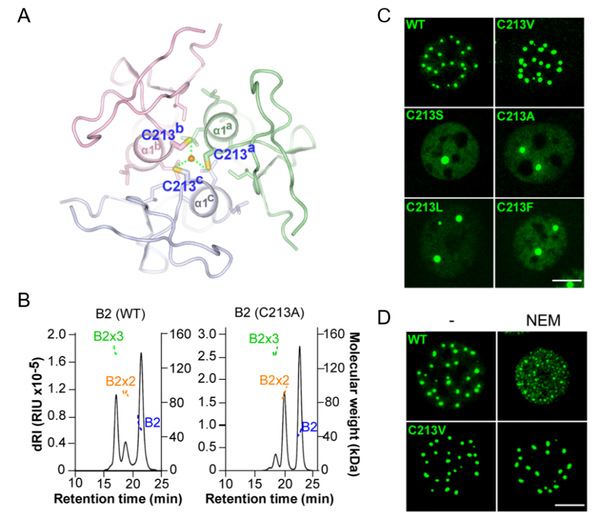
Figure 2. Arsenic docks on a C213 tri-cysteine pocket of B2 trimers
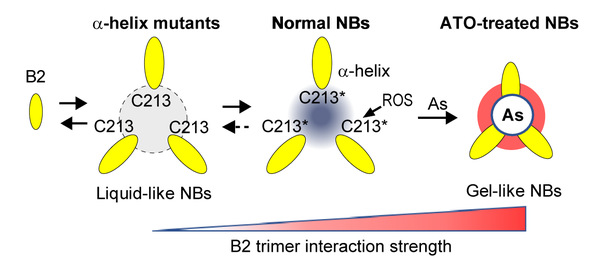
Figure 3. Model of PML NB liquid to gel-like transition controlled by the C213-containing B2 α1 helix hijacked by ATO
This study establishes that the PML B-box-2 structure reveals an alpha helix that drives B2 trimerization and positions a cysteine trio to form an ideal arsenic-binding pocket. Altering either of the latter impedes ATO-driven NB-assembly, PML sumoylation, and PML-RARA degradation, providing a mechanistic explanation for clinical ATO-resistance. The B2 trimer and the C213 trio create an oxidation-sensitive rheostat that controls PML NB assembly dynamics and downstream signaling in both the basal state and during stress response. These findings identify the structural basis for arsenic targeting of PML, which could pave the way for novel cancer drugs.
Source: The research team led by Professors Naranmandura Hua and ZHOU Chun